Los ácidos grasos poliinsaturados y sus derivados regulan infecciones respiratorias
Ruiz, Andy1,2; Romero-García, Aida Susana2; Mancilla-Jiménez, Raúl2; Juárez, Esmeralda1

2022, Número 1
2022; 81 (1)


Ruiz, Andy1,2; Romero-García, Aida Susana2; Mancilla-Jiménez, Raúl2; Juárez, Esmeralda1
RESUMEN
La regulación de la inflamación es un proceso fisiopatológico complejo que depende de la producción de lípidos oxigenados derivados de los ácidos grasos poliinsaturados esenciales, como el omega-3 y el omega-6, entre los que se encuentran las lipoxinas, resolvinas y protectinas, denominados mediadores lipídicos pro-resolvedores de la inflamación (SPM, del inglés specialized pro-resolving lipid mediators). La actividad de éstos se asocia con el control de procesos respiratorios infecciosos al modular la producción de citocinas proinflamatorias, evitar el daño por necrosis asociado a la inflamación, disminuir cargas microbianas y promover la regeneración de los tejidos. En este trabajo revisamos los aspectos bioquímicos, inmunológicos y fisiopatológicos de los SPM en la regulación de la inflamación en infecciones respiratorias.PALABRAS CLAVE
Ácido eicosapentaenoico, ácido docosahexaenoico, inflamación, infecciones respiratorias, mediadores lipídicos pro-resolvedores de la inflamación.Introducción
Los ácidos grasos poliinsaturados esenciales (PUFA, del inglés polyunsaturated fattty acids), como el omega-3, se obtienen de fuentes ricas de pescado, salmón, nueces y semillas de lino, mientras que las fuentes ricas en omega-6 incluyen aceites vegetales de maíz, cártamo, girasol, soja y algunos productos animales.1-3
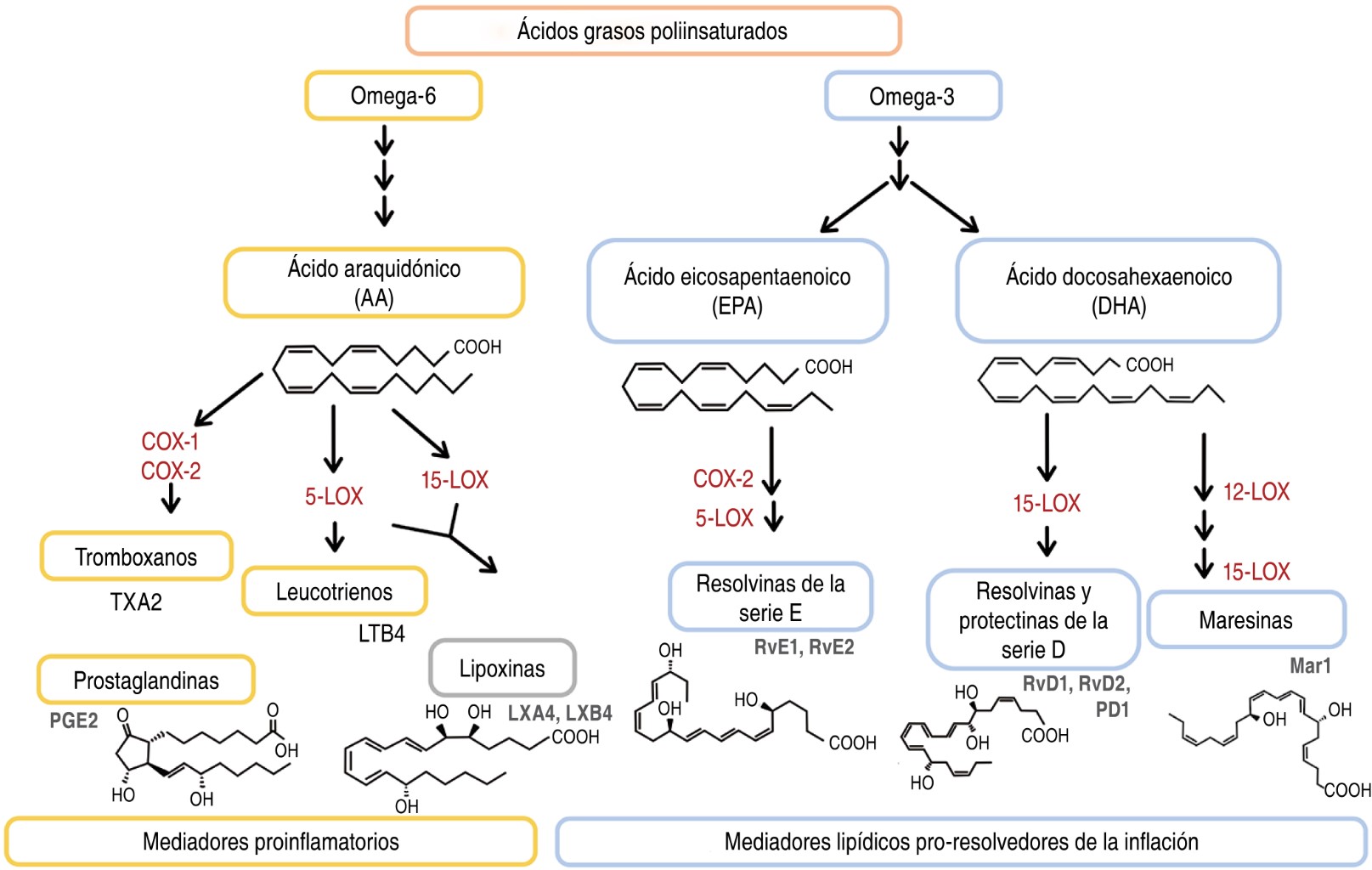
El estudio de los PUFA ha incrementado por su participación en la regulación de respuestas inflamatorias, como la producción de mediadores lipídicos pro-resolvedores de la inflamación (SPM, del inglés specialized pro-resolving lipid mediators). Los SPM derivados de los PUFA como el ácido linoleico (C18: Δ2, n-6), ácido araquidónico (AA, C20: Δ4, n-6), ácido eicosapentaenoico (EPA, 20:55,8,11,14,17) y ácido docosahexaenoico (DHA, 22:64,7,10,13,16,19) son generados a partir de reacciones enzimáticas mediadas por lipooxigenasas (LOX) y/o ciclooxigenasas (COX), que incluyen protectinas y resolvinas de la serie D, derivadas del DHA, resolvinas de la serie E, derivadas del EPA, y lipoxinas, derivadas del AA, como se muestra en la Figura 1.4-9
Tanto en modelos in vitro como in vivo, los SPM promueven el aclaramiento bacteriano al estimular la producción de péptidos antimicrobianos,7,10 incrementan la actividad fagocítica de los macrófagos11-13 y disminuyen la producción de citocinas proinflamatorias. Además, ayudan en la reparación tisular, aumentan las defensas del hospedero y mejoran su supervivencia.14,15
Se tiene evidencia de que las infecciones respiratorias se ven afectadas por el estado nutricional del paciente, el estado metabólico, la medicación, las complicaciones y el curso de la enfermedad pulmonar propiamente dicha,16-22 por lo que lograr el equilibrio entre los efectos protectores y perjudiciales de la respuesta inmune pueden ayudar a disminuir la morbimortalidad y las complicaciones en infecciones respiratorias. Por lo tanto, investigar los aspectos bioquímicos, inmunológicos y fisiopatológicos de los PUFA y sus derivados ayudarán a vislumbrar rutas, vías de administración y formulaciones nutricionales que ayuden a seleccionar estrategias para erradicar a los patógenos de vías respiratorias.
Papel fisiológico de los SPM
Las acciones biológicas de los SPM están mediadas por la activación de receptores afines. La señalización inicia localmente por los receptores asociados a proteínas G específicos (GPCR, por sus siglas en inglés G protein-coupled receptors) que se expresan en diferentes tipos de células (células polimorfonucleares [PMN], células dendríticas, monocitos, macrófagos, células epiteliales, fibroblastos, adipocitos, etcétera) y promueven selectividad tisular, ejerciendo su acción ante respuestas extracelulares. En la Tabla 1 se muestran algunos de los receptores SPM encontrados hasta el momento, así como sus agonistas, antagonistas y genes reguladores.23-30
ALX fue el primer receptor identificado, éste es activado por ligandos endógenos afines, incluidos la lipoxina A4 (LXA4) y las resolvinas D1 y D3 (RvD1 y RvD3), así como las desencadenadas por aspirina (AT-LXA4, AT-RvD1). RvD1 activa al receptor GPR32 que conduce a la regulación de varios micro-ARN (miRNA) involucrados en la orquestación de la inflamación aguda, incluidos miR-(miRNA)146b, miR-208a y miR-219. Este receptor también media las acciones biológicas de RvD5 en el contexto de infecciones bacterianas, por lo que su activación por RvD5 conduce a una mayor fagocitosis bacteriana en los macrófagos humanos y a una baja regulación de varios genes proinflamatorios, incluidos NF-κB (factor nuclear potenciador de las cadenas ligeras kappa de las células B activadas) y TNF-α (factor de necrosis tumoral alfa).31,32
El efecto biológico de las resolvinas está mediado por los receptores ALX, FPR2, DRV1, GPCR32, DRV2, GPCR18, ChemR23 o ERV1. Se ha demostrado que RvD1 inhibe el NF-κB canónico (p65/p50) y la activación de la vía NF-κB no canónica (p50/p50), lo que conduce a la inhibición de apoptosis y bloqueo en la producción de citocinas proinflamatorias, reduciendo la migración transendotelial de PMN, aumentando la actividad de los macrófagos, lo que resulta en la eliminación de células apoptóticas.33 Por otra parte, RvD1 es capaz de activar PPARγ (receptor de peroxisoma proliferador activado gamma) y suprimir la degradación de NF-κB vía p65.34
Algunos estudios han demostrado que RvD2 activa el receptor DRV2/GPCR18 controlando las funciones de fagocitos tanto en humanos como en ratones para dichos receptores, donde infecciones bacterianas fueron controladas, mejorando la supervivencia en murinos y brindando protección de órganos, mientras que estas acciones fueron mermadas en ratones transgénicos Knockout (KO) para DRV2.35
En el caso de RvE1, se ha demostrado que tiene una función como agonista para ChemR23/ERV y antagonista para el receptor LTB4 (BLT1) en PMN. Siendo capaz de inhibir el anión superóxido de PMN en respuesta al TNF-α o al péptido bacteriano N-formil-metionil-leucil-fenilalanina (f-MetLeuPhe), además estimula la fagocitosis de PMN apoptóticos por macrófagos. Mientras que en un modelo de periodontitis de conejo, la administración de RvE1 dio lugar a la regeneración de tejidos dañados, incluyendo hueso, en comparación con el uso de aspirina o esteroides como la dexametasona, inhibió selectivamente el tromboxano, demostrando su capacidad de ejercer efectos antiinflamatorios.26
Las evaluaciones de las concentraciones de los SPM en el organismo se realizan mediante técnicas de alta resolución estructural como cromatografía líquida-espectrometría de masas (LC-MS, del inglés liquid chromatography-mass spectrometry), metabololipidómica y espectroscopia UV. La información reportada a la fecha, sugiere que los niveles basales de los SPM están en los intervalos submicromolar y nanomolar.23,29,30,34,36,37 Shivakoti et al.38 realizaron un estudio comparativo de las concentraciones de algunos SPM, donde determinaron que pacientes australianos diabéticos (DM) presentaban concentraciones más altas de RvD1, RvD2, RvE1, RvE2 y Mar1 en suero, en comparación con pacientes con tuberculosis (TB) y pacientes con TB y diabetes (TB-DM), lo que indica que la infección promueve un desbalance entre estos mediadores lipídicos, dando pie a considerar los niveles de SPM como biomarcadores de enfermedad.
SPM en enfermedades respiratorias infecciosas y no infecciosas
El sistema respiratorio humano suele dividirse en vías respiratorias superiores y bajas. Las vías respiratorias superiores incluyen cavidades nasales, cavidad oral, senos paranasales, nasofaringe y laringe (que juegan un rol importante en la eliminación de partículas). Las vías respiratorias bajas incluyen la tráquea, bronquios principales, bronquios terminales y bronquios respiratorios, así como los alvéolos.39,40 Las infecciones pueden afectar a ambas vías respiratorias, siendo las más frecuentes la rinofaringitis aguda (resfriado común, ocasionado por rinovirus, coronavirus y el virus respiratorio sincitial [VRS], y más raramente por enterovirus, influenza y parainfluenza).41-47 En modelos murinos se ha demostrado que la infección por virus de la influenza H5N1 causa una desregulación en la expresión y señalización de SPM, tales como las lipoxinas,48 mientras que la administración exógena de PD1 logra inhibir la infección de este virus mejorando la supervivencia y función pulmonar.49 Por otra parte, Ramón et al.50 demostraron un efecto coadyuvante con la administración del ácido 17(S)-hidroxidocosahexaenoico (17-HDHA) tras la vacunación contra la influenza, al incrementar significativamente los niveles de anticuerpos anti-H1N1 en suero, así como el número de células B en médula ósea de murinos.
Otras de las infecciones frecuentes son la faringoamigdalitis (inflamación de las membranas orofaríngeas y amígdalas palatinas, ocasionada comúnmente por adenovirus, parainfluenza, virus de Epstein-Barr, Coxsackievirus y Streptococcus β-hemolítico del grupo A)43,44,51-53 y la rinosinusitis (inflamación de la mucosa que reviste los senos paranasales, ocasionada por Haemophilus influenzae, Staphylococcus aureus, Staphylococcus pyogenes, Bacteroides sp. y Fusobacterium sp.).51,54,55 En un modelo de infección con H. influenzae se ha encontrado que la administración de AT-RvD1 regula el transporte de leucocitos al pulmón, incrementando la fagocitosis de neutrófilos por macrófagos y reduciendo los niveles de la interleucina 6 (IL-6) y TNF-α.56
Por otra parte, la permeabilidad del epitelio alveolar puede desencadenar una respuesta inflamatoria por la entrada de diferentes agentes exógenos y endógenos que pueden estimular de forma persistente al organismo, lo que implica un reto para el mantenimiento de la homeostasis y la resolución de la inflamación.
Algunos microorganismos tienen la capacidad de establecerse de forma crónica como Mycobacterium tuberculosis, causante de TB, que maneja las cifras más altas de muertes por enfermedad infecciosa en todo el mundo después del virus de la inmunodeficiencia humana (VIH).57-59 En un modelo experimental de ratones deficientes en la 5-lipooxigenasa (5-LO, enzima encargada de la producción de lipoxinas), parece tener un mejor control de la infección por M. tuberculosis en comparación con ratones silvestres infectados con M. tuberculosis tratados con un inhibidor de la 5-LO, donde estos últimos tuvieron una mayor mortalidad y mayor carga bacteriana. Estos resultados sugieren que el control de la infección está relacionada con la producción de leucotrienos (vía proinflamatoria) más que por la producción de lipoxinas (vía antiinflamatoria).60 Mientras que en otro modelo in vitro de macrófagos humanos infectados con la cepa virulenta Mtb H37Rv tratados exógenamente con RvD1 y Mar1 se indujo la expresión de péptidos antimicrobianos como BPI (bactericidal permeability-increasing protein) y la catelicidina humana LL37, regulando la producción de TNF-α y controlando el crecimiento intracelular de Mtb.10 Estas investigaciones nos muestran estrategias que pueden llegar a utilizarse para apoyar el tratamiento actual de la TB, ya sea por suplementación de los precursores de los SPM como DHA/EPA o por administración exógena de los propios SPM.
Dentro de otros agentes externos que pueden causar afecciones respiratorias están los alérgenos (p. ej., las proteínas Derp2, presentes en las heces de los ácaros del polvo), partículas no degradables (como el asbesto) e incluso los cristales de partículas endógenas (p. ej., el colesterol),61-63 sin dejar de lado el humo del cigarro, asociado a enfermedades respiratorias crónicas, cardiovasculares y tumorales, que afectan la capacidad fagocítica de los macrófagos.39,64-68 Algunas investigaciones han demostrado que análogos de prostaglandinas y lipoxinas tienen propiedades físico-químicas que mejoran el uso de los glucocorticoides, ya que una disminución de estos últimos mejoran los efectos adversos, así como la resistencia a los esteroides en asma.69-71 Además, en un modelo de asma alérgica, se determinó que la administración de algunos activadores como TLR7 (toll like receptor 7) incrementaban los SPM derivados del DHA como PD-1, 17-HDHA y 14-HDHA ayudando al control de la eosinofilia caracterizada en este modelo animal, así como en otro modelo por administración intraperitoneal de RvE1.72,73
La enfermedad pulmonar obstructiva crónica (EPOC), el síndrome de dificultad respiratoria neonatal (SDRN), el síndrome de dificultad respiratoria aguda (SDRA), la lesión pulmonar aguda (ALI, acute lung injury) y el asma, son afecciones del sistema respiratorio que presentan altas incidencias, morbilidad y mortalidad. La EPOC es caracterizada por la limitación del flujo aéreo y se asocia a una respuesta inflamatoria anormal de los pulmones a partículas nocivas o a gases. El humo del tabaco es el principal factor de riesgo,74-76 le sigue la contaminación del aire,77,78 la exposición laboral a polvos y productos químicos, la recurrencia de infecciones respiratorias durante la niñez o la predisposición genética. Algunos estudios en modelos murinos se han enfocado en la administración exógena de LXA4, ya que este SPM compite con las proteínas amiloides séricas A (SAA, Serum amyloid A) por los GPCR FPRR/ALX, SAA incrementa considerablemente en infecciones y está relacionado en el reclutamiento excesivo de neutrófilos en EPOC, por lo tanto, ambos actúan como antagonistas, lo que puede ayudar a prevenir la inflamación crónica y el enfisema pulmonar.75,76,79
Por otra parte, el SDRN, el SDRA y ALI son enfermedades relacionadas con el sistema surfactante pulmonar, pero también ocurren con mayor frecuencia en el contexto de neumonía, sepsis, aspiración de contenido gástrico o trauma severo, a diferencia del asma, considerada como un trastorno inflamatorio heterogéneo altamente prevalente de las vías respiratorias por inflamación ocasionada por diversos alérgenos.80-82 Eickmeier et al.83 encontraron que la administración de AT-RvD1 en un modelo murino de ALI disminuyó la cantidad de neutrófilos del líquido de lavado broncoalveolar, mejoró la integridad de la barrera epitelial y endotelial, disminuyó significativamente los niveles de citocinas proinflamatorias como la interleucina 1β (IL-1β), IL-6 y TNF-α, así como la translocación nuclear de p65 fosforilada por NF-κB, por lo que este SPM podría funcionar también para el SDRN y SDRA.
En la actualidad, la enfermedad de COVID-19 ocasionada por el coronavirus SARS-CoV-2, ha impulsado la búsqueda de nuevas estrategias terapéuticas para combatir la gravedad de la enfermedad, enfocándose en la eliminación de las respuestas exacerbadas por producción de citocinas proinflamatorias,84 donde algunos grupos se enfocaron en los precursores de los SPM, como la suplementación del PUFA omega-3, encontrando mejoras en algunos parámetros de la función respiratoria y renal de pacientes críticos con COVID-19 evaluados durante un mes, en comparación con pacientes sin suplementación.85 La evaluación de los niveles de algunos SPM en pacientes diagnosticados con COVID-19 mostró que pacientes críticos presentaban concentraciones más bajas de los SPM que quienes fueron dados de alta.86,87 Recchiuti et al.88 encontraron en un modelo in vitro de macrófagos con o sin fibrosis quística expuestos a la glicoproteína S (Spike 1) del virión SARS-CoV-2, que tanto RvD1 como RvD2 fueron capaces de regular las funciones inflamatorias al modular miR-16, miR-29a y miR-103, y simultáneamente incrementaron selectivamente miR-223 y miR-125a, involucrados en la activación de NF-κB y la polarización inflamatoria de los macrófagos. No obstante, queda pendiente dilucidar si los diferentes factores de riesgo asociados a la enfermedad que incluyen edad avanzada, hipertensión, diabetes, obesidad u otras comorbilidades tienen alguna asociación con los SPM.
Como se ha visto a lo largo del texto, el análisis de los efectos biológicos de los SPM en las infecciones respiratorias puede dar lugar a nuevas propuestas de inmunomodulación terapéutica. De manera reciente, De Toledo y colaboradores64 demostraron que la fracción 39 del moco de la babosa Phyllocaulis boraceiensis contiene PUFA con potente actividad antiviral contra el virus del sarampión y el virus de la influenza. La viabilidad celular y la toxicidad del moco se evaluaron en células de riñón canino Madin-Darby (MDCK) mediante el ensayo de Bromuro de 3-(4,5- dimetiltiazol-2-ilo)-2,5-difeniltetrazol (MTT), donde demostraron que PUFA hidroxilados interferían con la unión del virus de la influenza al receptor de la célula huésped, provocando reducción en los títulos virales.
Por otra parte, en un modelo in vitro de neutrófilos humanos, la administración de lipoxina desencadenada por aspirina (15-epi-LXA4) anula la supresión de la apoptosis de neutrófilos con mieloperoxidasa (MPO, enzima con actividad microbicida) al bloquear la señalización mediada por la integrina β2, mejorando la resolución de la lesión pulmonar sostenida por MPO.89-91 Mientras tanto, en un modelo murino se evaluó la lesión pulmonar aguda por inyección intraperitoneal de Escherichia coli en ratones y se encontró que el tratamiento posterior con 15-epi-LXA4 promovió la apoptosis de neutrófilos y mejoró la resolución de la inflamación en la lesión pulmonar, comparable a ratones tratados con RvD1 previo a ALI por LPS, donde la RvD1 mejora la tasa de supervivencia de ratones expuestos a ALI con inhibición de TNF-α, IL-6 y disminución de la expresión de COX-2.92 Resultados similares se han encontrado con la administración de RvE1 en un modelo murino de neumonía, con exposición a E. coli, donde hubo reducción en la producción de citocinas proinflamatorias, disminución de PMN y reducción de las cargas bacterianas de E. coli, mejorando la supervivencia de los murinos.11
Por otro lado, Raposo et al.93 evaluaron la ingesta nutricional de vitamina C, vitamina E, DHA, AA, selenio y zinc, en una cohorte con más de 1,500 individuos de entre 25 a 64 años que fueron seguidos durante nueve meses, encontraron una asociación en la susceptibilidad a la infección del tracto respiratorio superior en mujeres que en hombres por disminución en el consumo de DHA, AA y vitaminas C y E. En contraposición, en fibroblastos pulmonares humanos y línea celular bronquial (BEAS-2B) se ha demostrado que los PUFA como el DHA, EPA y ALA (α-linolénico) provocan una amplificación de las respuestas inflamatorias a los componentes virales y bacterianos, con producción de IL-6 y CXCL8, lo que sugiere que los ácidos grasos poliinsaturados no tienen efectos antiinflamatorios en estas células pulmonares.94 Un breve resumen de la acción de los SPM se muestran en la Tabla 2.
Participación de los SPM en la resolución de inflamación
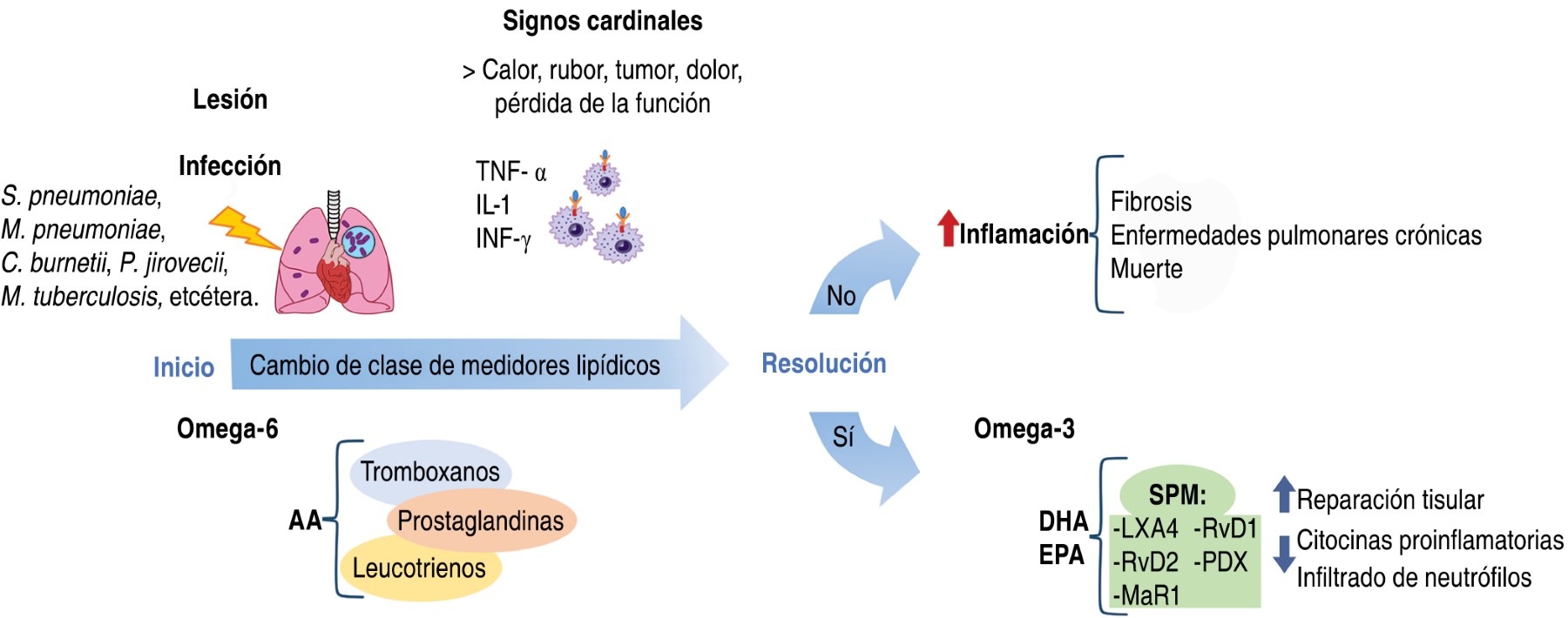
La inflamación es una respuesta del sistema inmunológico de un organismo al daño ocasionado por patógenos o por sustancias de naturaleza biológica, química, física o mecánica y, dependiendo de la duración, puede clasificarse en aguda o crónica.
La inflamación aguda comprende cambios significativos en niveles plasmáticos de histamina, bradiquinina, prostaglandinas, leucotrienos, tromboxanos y citocinas proinflamatorias (TNF-α, IL-1, IL-1β, IL-2 e IL-6), cruciales para controlar y eliminar a agentes nocivos,99-104 pero si la inflamación aguda se mantiene, conduce a inflamación crónica con repercusiones sistémicas y deletéreas para el hospedero, como infiltración tisular por agregados de leucocitos (formación de granulomas), biosíntesis descontrolada de colágeno, dando lugar a fibrosis o cirrosis, pérdida permanente de la función del tejido normal (functio laesa) o lesiones oxidativas en el ácido desoxirribonucleico (ADN), dando lugar a enfermedades degenerativas como enfermedades autoinmunes, trastornos cardiovasculares, osteoporosis, artritis reumatoide, enfermedad de Alzheimer, ciertos tipos de cáncer e incluso la muerte.103
Por lo que es perentorio la participación de los SPM en el mantenimiento y la respuesta de la inflamación, realizando el cambio de un fenotipo proinflamatorio a uno antiinflamatorio, ayudando así a la reparación del tejido y al restablecimiento de la homeostasis,105 como se muestra en la Figura 2.104
Conclusiones
Los PUFA y sus derivados, los SPM, tienen un efecto protector y de control en la eliminación de microorganismos patógenos, inflamación y reparación tisular, lo que los hace candidatos importantes para la búsqueda de nuevas estrategias terapéuticas, así como posibles biomarcadores. Es necesario profundizar en el conocimiento de sus mecanismos de señalización, vías de síntesis, producción de sus epímeros e investigaciones que evalúen el consumo de PUFA y los niveles de SPM en sujetos sanos respecto a pacientes con enfermedades respiratorias para comprender mejor la relación entre los perfiles generales de PUFA en la dieta y su impacto en futuras intervenciones nutricionales o farmacológicas como estrategias para erradicar a patógenos de diversas afecciones respiratorias.
Agradecimientos
Andy Ruiz agradece al Programa de Doctorado en Ciencias Biológicas de la UNAM, así como al Consejo Nacional de Ciencia y Tecnología (CONACyT) por la beca recibida durante sus estudios y agradece los comentarios realizados por el Dr. José Ángel Santiago Terrones.
AFILIACIONES
1Instituto Nacional de Enfermedades Respiratorias Ismael Cosío Villegas, CDMX, México; 2Universidad Nacional Autónoma de México, CDMX, México.Conflicto de intereses: Los autores declaran no tener conflicto de intereses.
REFERENCIAS
Tvrzicka E, Kremmyda LS, Stankova B, Zak A. Fatty acids as biocompounds: Their role in human metabolism, health and disease - a review. part 1: Classification, dietary sources and biological functions. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2011;155(2):117-130. Available in: https://doi.org/10.5507/bp.2011.038
Cederholm T, Jensen GL, Correia MITD, Gonzalez MC, Fukushima R, Higashiguchi T, et al.; and GLIM Core Leadership Committee, GLIM Working Group. GLIM criteria for the diagnosis of malnutrition – A consensus report from the global clinical nutrition community. J Cachexia, Sarcopenia Muscle. 2019;10(1):207-217. Available in: https://doi.org/10.1002/jcsm.12383
Polus A, Zapala B, Razny U, Gielicz A, Kiec-Wilk B, Malczewska-Malec M, et al. Omega-3 fatty acid supplementation influences the whole blood transcriptome in women with obesity, associated with pro-resolving lipid mediator production. Biochim Biophys Acta. 2016;1861(11):1746-1755. Available in: https://doi.org/10.1016/j.bbalip.2016.08.005
Guzmán-Beltrán S, Carreto-Binaghi LE, Carranza C, Torres M, Gonzalez Y, Muñoz-Torrico M, et al. Oxidative stress and inflammatory mediators in exhaled breath condensate of patients with pulmonary tuberculosis. a pilot study with a biomarker perspective. Antioxidants. 2021;10(10):1572. doi: 10.3390/antiox10101572.
Abdillahi SM, Tati R, Nordin SL, Baumgarten M, Hallgren O, Bjermer L, et al. The pulmonary extracellular matrix is a bactericidal barrier against Haemophilus influenzae in chronic obstructive pulmonary disease (COPD): Implications for an in vivo innate host defense function of collagen VI. Front Immunol. 2018;9:1988. doi: 10.3389/fimmu.2018.01988.
Doaei S, Gholami S, Rastgoo S, Gholamalizadeh M, Bourbour F, Bagheri SE, et al. The effect of omega-3 fatty acid supplementation on clinical and biochemical parameters of critically ill patients with COVID-19: a randomized clinical trial. J Transl Med. 2021;19(1):128. doi: 10.1186/s12967-021-02795-5.