Los eicosanoides como reguladores de procesos inflamatorios e inmunológicos en la tuberculosis pulmonar
Escalona-Sarabia, Ana Luisa1; Juárez, Esmeralda2

2022, Número 2
2022; 81 (2)


Escalona-Sarabia, Ana Luisa1; Juárez, Esmeralda2
RESUMEN
La tuberculosis (TB) humana es un problema de salud pública. Aunque es una enfermedad curable, el protocolo de tratamiento actual es largo y difícil de mantener. Debido al limitado avance de nuevos tratamientos en los últimos años, las terapias dirigidas al hospedero han tomado un mayor interés, siendo una de las propuestas la intervención en la ruta metabólica de los eicosanoides, los cuales son mediadores químicos locales que favorecen o limitan la inflamación. Durante la TB, un estado de hiperinflamación genera daño al parénquima deteriorando la función respiratoria. A pesar de la importancia de este circuito, los reportes sobre su utilidad en tuberculosis en ocasiones son controversiales y no concluyentes. Con el objetivo de conocer e integrar la información publicada, en este trabajo se analizaron diversos estudios que buscan definir el papel de los eicosanoides en la infección por M. tuberculosis. Para ello, analizamos el papel de los eicosanoides posinfección in vivo o in vitro, y las intervenciones terapéuticas en su ruta metabólica in vivo pre- o posinfección. Además, proponemos un algoritmo que permita para optimizar futuras investigaciones sobre eicosanoides y su utilización como blancos terapéuticos de la TB.PALABRAS CLAVE
tuberculosis, eicosanoides, terapias dirigidas al hospedero.Introducción
La tuberculosis (TB) humana, ocasionada por Mycobacterium tuberculosis, es una de las 10 principales causas de muerte en el mundo.1 En México se reportan más de 20,000 casos nuevos por año, mayoritariamente la forma pulmonar.2,3 La inflamación crónica que se presenta en los pulmones causa daño al parénquima y deterioro de la función respiratoria,4,5 convirtiendo al circuito inflamatorio un blanco terapéutico para preservar la función pulmonar. Así, las terapias dirigidas al hospedero, como moduladores de citocinas (IFN-γ y TNF-α), antifibróticos, o moléculas que activan macrófagos son ideales por su capacidad de modular al sistema inmune y limitar el daño tisular posinfección.6,7
Intervenir la ruta metabólica de los eicosanoides es una opción terapéutica para prevenir la inflamación y preservar la función pulmonar. Los eicosanoides son lípidos derivados de ácidos grasos poliinsaturados (PUFA, por sus siglas en inglés) que se obtienen de la ingesta de ácidos grasos omega-6 o ω-6 (ácido araquidónico [AA]) y omega-3 o ω-3 (ácido eicosapentaenoico [EPA] y ácido docosahexaenoico [DHA]). Estos lípidos son sintetizados enzimáticamente por lipoxigenasas (LOX) y/o ciclooxigenasas (COX) en leucocitos, células endoteliales y plaquetas.7 Existen dos familias principales de los PUFA con actividades antagónicas, los proinflamatorios y los prorresolutorios.
Los eicosanoides proinflamatorios incluyen a los tromboxanos, prostaglandinas y prostaciclinas sintetizados por la COX-2, y a los leucotrienos, sintetizados por las 5 y 15-LOX a partir del AA. Los eicosanoides proinflamatorios son quimioatrayentes de neutrófilos, activan la fagocitosis en macrófagos alveolares,8 median el tráfico de células,9 e inducen necrosis, edema, aumento del flujo sanguíneo y producción de citocinas proinflamatorias. Los eicosanoides prorresolutorios, o mediadores lipídicos especializados en resolución de la inflamación (SPM por sus siglas en inglés), son sintetizados por las 5 y 15-LOX e incluyen las lipoxinas, resolvinas, protectinas y maresinas.10 Estos lípidos limitan el influjo de neutrófilos, bloquean la producción de especies reactivas de oxígeno, inducen apoptosis y aumentan la actividad fagocítica de los macrófagos favoreciendo el regreso a la homeostasis y la regeneración celular.11,12
A pesar de lo importante que este circuito es para las infecciones respiratorias, su contribución en la regulación de la inflamación en TB requiere ser revisada en función de los modelos experimentales publicados. En esta revisión, consideraremos los mecanismos moleculares de los eicosanoides involucrados en el proceso de regulación de la inflamación durante la TB en modelos animales; observaremos que algunos fenómenos que se presentan en modelos animales no se pueden replicar en humanos y viceversa; y evaluaremos la validez de utilizar terapias dirigidas al hospedero en función de los mecanismos moleculares que los eicosanoides regulan.
Modelos experimentales utilizados para estudiar el papel que los eicosanoides juegan en la tuberculosis
Para definir el papel que los eicosanoides desempeñan en la infección por M. tuberculosis establecimos tres diseños experimentales: 1) cuantificación de eicosanoides posinfección; 2) intervención terapéutica realizada in vivo con ácidos grasos poliinsaturados (PUFA) antes de la infección; y 3) intervención terapéutica con PUFA posinfección.
Cuantificación de eicosanoides posinfección. Modelos con animales infectados in vivo con cepas virulentas de M. tuberculosis (H37Rv, Erdman o HN878), así como muestras de pacientes, revelan cambios en los niveles de eicosanoides durante y después de la infección con TB que correlacionan con la severidad de la enfermedad.13,14 Los eicosanoides principalmente estudiados son los derivados del AA: PGE2, lipoxina A4 (LXA4) y LTB4. La presencia de LXA4 y LTB4 está asociada a necrosis y daño tisular,12 mientras que la presencia de PGE2, o su receptor EP2, tienen efectos opuestos y se asocian a resistencia a la infección.12-14 El aumento de LXA4 se asocia con una mayor susceptibilidad a la enfermedad y PGE2 con una respuesta protectora. En el tejido pulmonar de pacientes con TB pulmonar se observa la presencia de AA y LTA4H (leucotrieno A4 hidrolasa) en los centros necróticos y la presencia de ciclooxigenasa en la periferia de las lesiones.15 Los pacientes con TB pulmonar, TB multidrogorresistente (MDR) o TB latente tienen mayores cantidades circulantes de PGE2, LTB4 y LXA4 que las personas sanas.16-18
Aunque en algunos estudios se midieron otros eicosanoides, como maresina (Mar1, Mar2), resolvinas RvD (RvD1-6) y RvE (RvE1-4); prostaglandinas (PGF2, PGD2), protectinas (PD1) y precursores de eicosanoides 12-HETE o 15-HETE,19-21 no se encontró relevancia de éstos en TB individualmente. Más bien, los efectos de los eicosanoides dependen de su contribución relativa.22 Por ejemplo, un aumento en la proporción de LTB4/Mar1 en suero distingue a pacientes de individuos sanos,19 la proporción LTB4/LXA4 disminuye postratamiento,13 las conexiones entre SPM y lípidos proinflamatorios son mayores en pacientes con TB-DM en comparación con pacientes con sólo TB,20 y los niveles de PGE2 dependen de la proporción entre LTB4/LXA4.23
La relación entre PGE2 y LXA4 es antagónica en cultivos de macrófagos, LXA4 induce necrosis y PGE2 induce apoptosis para protección celular ante la infección.24-26 Al igual que en las determinaciones realizadas in vivo, el aumento de LXA4 está asociado con una mayor susceptibilidad a la infección, mayor grado de inflamación y carga bacilar. Además, la infección por M. tuberculosis aumenta la liberación de AA en macrófagos y su transformación por la 5-LOX produciendo LXA4.24 La inhibición de la síntesis de LXA4 protege contra la necrosis,12 y por lo mismo, su inducción parece ser una estrategia de supervivencia de la micobacteria. Respecto a los SPM, RvD1 y Mar1, éstos inducen mecanismos antiinflamatorios y restauran la síntesis de péptidos antimicrobianos en macrófagos humanos infectados con la cepa virulenta M. tuberculosis H37Rv.25 La falta de medición de otros eicosanoides impide conocer el impacto real del circuito inflamación-resolución durante la infección por M. tuberculosis.
Intervenciones terapéuticas realizadas in vivo antes de la infección. Las investigaciones en las que se administran eicosanoides o se aplican terapéuticos que modifican sus rutas metabólicas antes de la infección permiten explicar los mecanismos involucrados durante el proceso de enfermedad. Las intervenciones más comunes son el uso de dietas enriquecidas con ácidos grasos omega-3 y omega-6, dietas deficientes en esos ácidos grasos, y fármacos inhibidores de la síntesis de eicosanoides.
Ratones tratados con dietas suplementadas con ω-626,27 y cobayos alimentados con suplementos de ω-328 presentan un aumento en la carga bacteriana, pero ratones BALB/c y C3HeB/FeJ alimentados con dietas enriquecidas en omega-3 muestran reducción de la carga bacteriana y de la cantidad de citocinas proinflamatorias liberadas al ambiente local.26,29,30 En humanos, un estudio longitudinal-prospectivo reveló que había mayor riesgo a desarrollar TB a mayor consumo de colesterol y un menor riesgo a desarrollar TB a mayor consumo de ω-3 y ω-6 de origen marino.31 Sin embargo, debido a sus efectos biológicos antagónicos, es difícil llegar a una conclusión cuando el consumo es variado y ocurre antes de la infección.
La intervención medicamentosa incluye inhibidores de 5-LOX o COX-2 para manipular la ruta metabólica. Inhibir la 5-LOX tiene efectos negativos porque aumenta la susceptibilidad a la infección disminuyendo la cantidad de leucocitos y aumentando la carga bacteriana.32-34 El mecanismo molecular implicado es incierto, pues la 5-LOX participa en la síntesis de LTB4 y LXA4 a partir del AA, ambos con efectos antagónicos, y también en la producción de resolvinas a partir de DHA y EPA, las cuales son prorresolutorias. En los casos en que se administró inhibidor de 5-LOX, la ausencia de resolvinas, y no la de LTB4, podría ser la razón de la falta de control de la infección.
La inhibición de la COX-2 tiene beneficios como disminución de carga bacteriana, el tamaño y presencia de granulomas y la mortalidad.19,32-35 La inhibición de la COX-2 implica el bloqueo de prostaglandinas, prostaciclinas y tromboxanos, cuyos efectos biológicos en TB no han sido explorados. Esta estrategia tiene potencial profiláctico para personas expuestas por contacto cercano a los pacientes. Sin embargo, en la mayoría de estos ensayos, los tratamientos se administraron antes de la infección y se continuaron durante la infección, por lo que es difícil saber si el resultado se debe a la activación de mecanismos previos a la infección o al sostenimiento de los mismos posinfección.
Intervenciones terapéuticas realizadas posinfección. Las intervenciones realizadas posinfección incluyen suplementación directa de eicosanoides (Tabla 1) o inhibición farmacológica de su síntesis (Tabla 2). Los esquemas que utilizan cambios en la dieta o agonistas de PGE2, LTB4 u otros eicosanoides posinfección son escasos. En general, la suplementación directa con eicosanoides reduce la producción de citocinas proinflamatorias, influye en la patogenicidad19 o confiere protección.36,37 Paradójicamente, PGE2 es un proinflamatorio con efectos inmunosupresores.12 Además, RvD1 y Mar1 tienen efectos antiinflamatorios sin detrimento de los mecanismos antimicrobianos.25
Por otro lado, durante la infección se producen eicosanoides de ambos tipos,38-40 y las dietas con suplementos de DHA/EPA tienen efectos antiinflamatorios,41,42 pero la relación entre ambos fenómenos se desconoce. Se requieren más investigaciones para conocer el verdadero potencial de la suplementación con DHA/EPA durante la TB en humanos.
La inhibición farmacológica de la síntesis de eicosanoides durante la infección por M. tuberculosis (Tabla 2) descansa principalmente en el uso in vivo, in vitro y ex vivo de inhibidores de la COX-2 (ácido niflúmico [AN], aspirina, celecoxib, etoricoxib o ibuprofeno). Esta inhibición generalmente causa reducción de la inflamación por disminución de la producción de citocinas, reducción del daño tisular y aumento en la sobrevida. Controversialmente, inhibir la COX-2 puede amentar la carga bacteriana cuando el tratamiento se aplica en fases tempranas de la infección,43 probablemente debido a los efectos inmunosupresores de PGE2,12 mientras que el uso tardío de estos inhibidores permite una mejor resolución de la enfermedad, con reducción de la carga bacteriana y la inflamación, lo que protege contra el daño tisular.33,44-47 Es necesario tomar con precaución la inhibición de la COX-2, pues sus efectos inmunosupresores pueden afectar al paciente durante las primeras fases de la enfermedad y se requieren más estudios para determinar su eficacia en TB.
Mecanismos moleculares asociados al metabolismo de los eicosanoides en la infección por M. tuberculosis
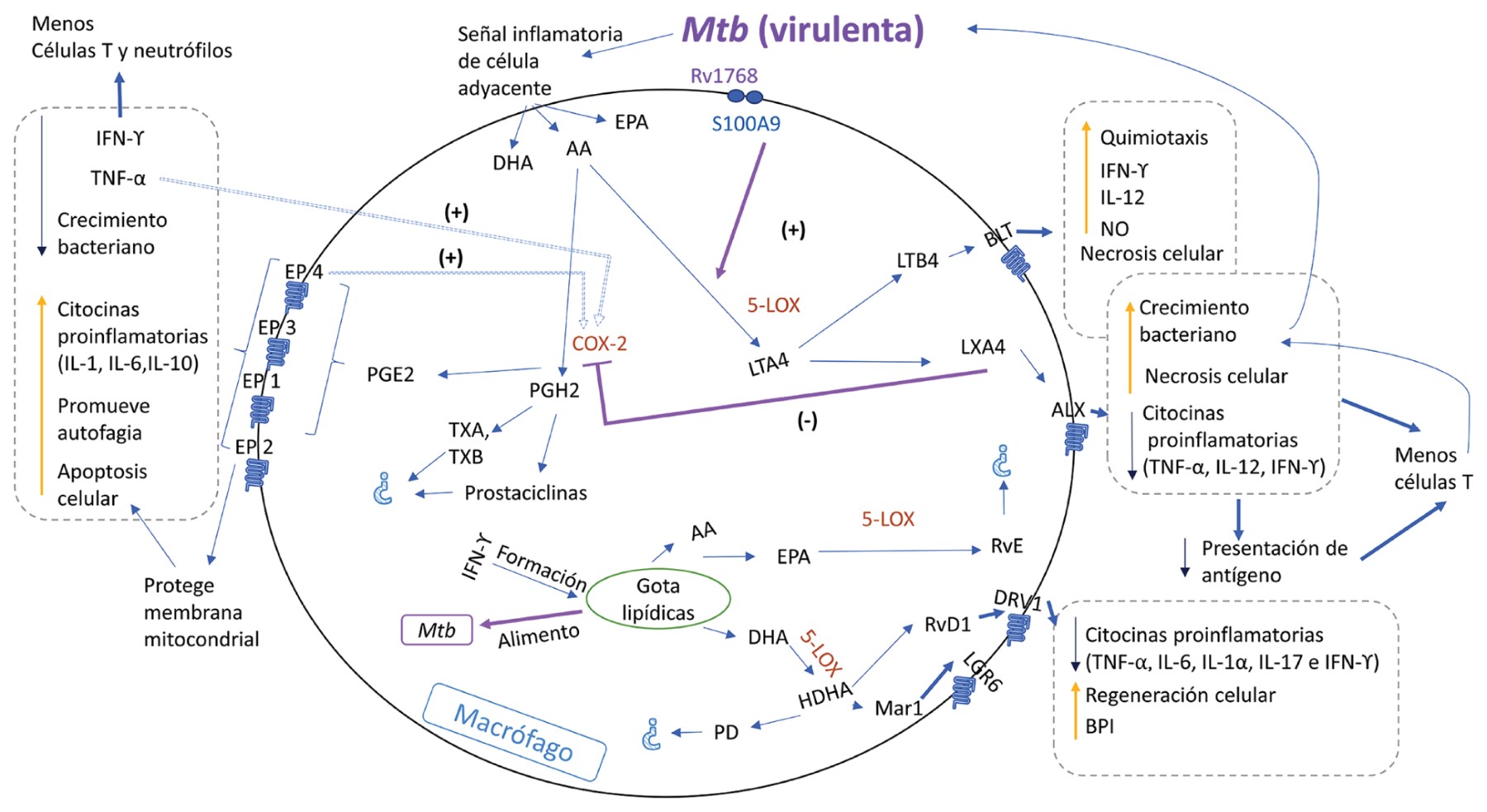
Durante las primeras horas de infección, el AA de la membrana nuclear y plasmática es procesado por las COX-1 y COX-2 en PGH2, la cual es convertida a PGE2 por cPGES, mPGES-1 o PGES-2.48 En macrófagos infectados, la producción de PGE2 se correlaciona con disminución de la fagocitosis, producción de óxido nítrico, prevención de necrosis, aumento de citocinas proinflamatorias e inducción de apoptosis, protegiendo la membrana mitocondrial, promoviendo la reparación de la membrana plasmática y aumentando el control de la inmunidad innata ante la infección micobacteriana.14,18,49 PGE2 también activa la autofagia potenciando la eliminación de la bacteria en autofagolisosomas. La autofagia, por su parte, controla la inflamación mediante regulación de señalizaciones de la inmunidad innata, modulación de la secreción de mediadores inmunes y eliminación de agonistas endógenos del inflamasoma.50
Después de las primeras 24 horas postinfección, aumenta la producción de LXA437 en macrófagos, la cual provoca un cambio en el metabolismo de AA a cargo de la 5-LOX.24 El aumento en los niveles de LXA4 está asociado con reducción de necrosis, carga bacilar, citocinas proinflamatorias, permeabilidad vascular, entrada de leucocitos polimorfonucleares (PMN) a los sitios de la inflamación y de la respuesta protectora tipo Th1.14,15,18
El efecto de la 5-LOX sobre el AA también provoca un aumento de LTB4. Durante las primeras fases de inflamación, sólo las células mesoteliales y macrófagos son capaces de liberar LTB4 al espacio pleural en respuesta de un estímulo de inflamación inicial. Establecido el proceso inflamatorio, otras células, como los neutrófilos, producen LTB4 amplificando el proceso inflamatorio.51 LTB4, a su vez, induce necrosis, aumento en la producción de óxido nítrico y quimiotaxis (Figura 1).
Por otro lado, el AA también se metaboliza en tromboxanos y prostaciclinas,19,52 pero éstos no se han asociado a TB. Otros PUFA de la membrana que se metabolizan durante la inflamación y estrés celular son el DHA y EPA. La conversión de precursores omega-6 y omega-3 en PUFA es controlada por enzimas desaturasas de ácidos grasos (FADS) 1 y 2.36 Posteriormente, éstos son transformados por la 5-LOX en maresina, protectinas y resolvinas. Aunque han sido reportados durante la TB,35,53,54 se desconoce la participación de estos eicosanoides en el proceso de la enfermedad, a excepción de resolvina D1 (RvD1) y maresina 1 (Mar1) que contribuyen al control de la infección por M. tuberculosis in vitro, mediante el aumento de la proteína bactericida de aumento de permeabilidad (BPI) y regeneración celular.25
Alcances de las terapias de intervención de la ruta metabólica de los eicosanoides
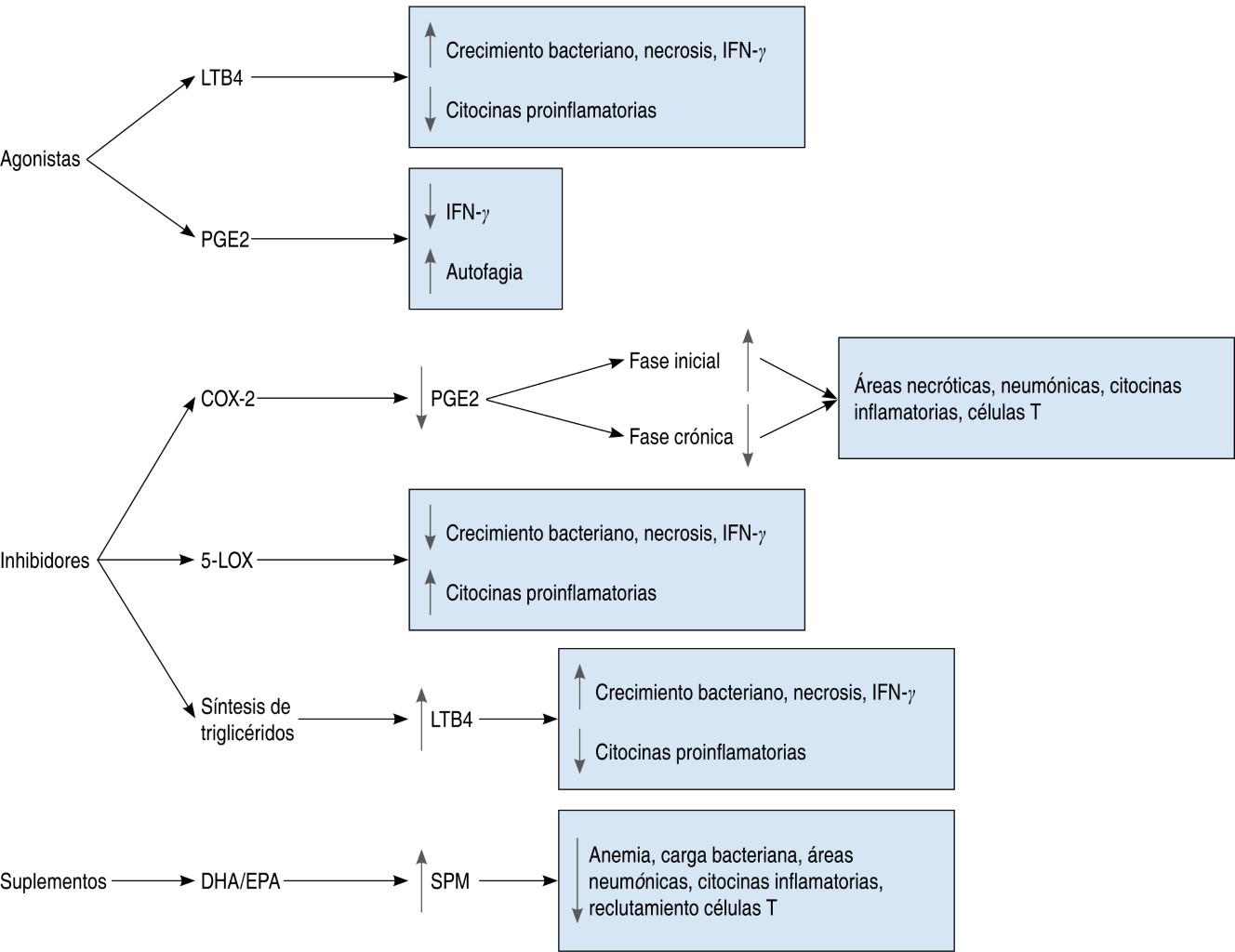
Las intervenciones realizadas posinfección han permitido entender los alcances de estas terapias dirigidas al hospedero (Figura 2). Actualmente se han inhibido tres rutas metabólicas involucradas en la síntesis de eicosanoides: COX-2, 5-LOX y la síntesis de triglicéridos. Los inhibidores de COX-2 disminuyen la producción de PGE2 provocando efectos diferentes dependiendo del tiempo en el que son administrados;43 al inicio de la enfermedad, inducen un aumento de las áreas necróticas, una mayor cantidad de citocinas inflamatorias y de células T;37 en cambio, la inhibición de COX-2 en etapas crónicas disminuye las lesiones y la severidad de la enfermedad.36 En general, no se encontraron interferencias con el uso de los antibióticos convencionales.
La inhibición de 5-LOX reduce el crecimiento bacteriano y la necrosis, pero aumenta las citocinas proinflamatorias;23 se desconoce si este efecto se debe al aumento de LXA4 o a la reducción de LTB4. La síntesis de triglicéridos regula a la alza la 5-LOX, pero la inhibición de éstos reduce la carga bacteriana, posiblemente porque representa disminución de nutrientes.38,40 Con el uso de suplementos alimenticios DHA/EPA, modelos animales revelan una reducción en anemia, inflamación y la carga bacteriana y aumento de la síntesis de SPM.54 Se desconoce el impacto que esta intervención tendría en humanos; sin embargo, se sabe que SPM derivados del DHA/EPA, MAR1 y RvD1 en cultivos in vitro de células humanas tienen efectos de regeneración celular y disminución de la inflamación.25 Sin embargo, estudios sobre polimorfismos genéticos reportan una mayor susceptibilidad a la enfermedad en poblaciones con variaciones en genes de 5-LOX,8 LTA4H55 o EP2;56 mientras que en otros no se reportan asociaciones entre esos mismos genes y la gravedad de enfermedad.57-59
Aunque los modelos animales ofrecen una variedad de recursos para el estudio de la tuberculosis, algunas limitaciones de los modelos experimentales utilizados actualmente impiden el análisis crítico y la implementación de intervenciones terapéuticas en humanos. Por ejemplo, no todas las cepas de ratón experimentan una respuesta predominantemente inflamatoria, mientras que humanos inmunocompetentes experimentan exacerbación de la inflamación.35 La variedad de cepas de ratón utilizadas influye en el resultado; en muchos casos fue necesario modificar algún gen que permitiera el estudio de la ruta metabólica de interés.60 Los ratones C3HeB/FeJ y Sv129 son extremadamente susceptibles a la infección por M. tuberculosis y los C57BL/6J y BALB/c son resistentes.61-63 En ratones, la acción biológica de PGE2 es mediada por cuatro proteínas ligadas a receptores de prostanoides EP1, EP2, EP3 y EP4. Estos receptores también son expresados en macrófagos humanos.14 No obstante, en macrófagos murinos infectados hay mayor cantidad de EP4 en comparación con EP2, lo que no ocurre en macrófagos humanos.64
En humanos, la forma más frecuente es la tuberculosis pulmonar; sin embargo, en investigación se estudia principalmente el plasma y células sanguíneas de donadores, que no reflejan lo que ocurre en el espacio alveolar. Actualmente, distintos tipos de tomografías se han utilizado para el monitoreo de la historia natural de la enfermedad, pero es difícil poder llevar a cabo estudios experimentales en humanos.65
Otros modelos animales se han utilizado. Por ejemplo, conejos infectados con M. tuberculosis HN878 producen lesiones parecidas a las encontradas en humanos. Los patrones de distribución de AA dentro del granuloma son similares en humanos y conejos,15 lo que haría del conejo un mejor modelo; sin embargo, en conejos normalmente se utilizan cepas M. bovis para el estudio de tuberculosis,65 los conejos son más caros de mantener y su gran susceptibilidad al estrés demanda un control estricto de los factores ambientales. Además, las distintas respuestas biológicas entre razas y las posturas de las distintas asociaciones de defensa de los animales impiden que se tenga más información.66
En cuanto a la suplementación con ω-3 (omega-3), los modelos experimentales son muy diversos y no predicen con el resultado esperado en humanos. Inclusive, las dosis experimentales utilizadas no representan de manera realista el aporte de la dieta en humanos;67 la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) recomienda una ingesta diaria de 250 mg de EPA + DHA.68 En los últimos años, la demanda de suplementos ha ido en aumento, pero se desconoce la cantidad necesaria de consumo de cada uno de ellos por separado, ya que cada uno tiene un metabolismo diferente.69 En México, también se desconoce el consumo promedio de DHA/EPA; sin embargo, a raíz de la pandemia de COVID-19, el consumo de pescado (fuente principal de estos ácidos grasos) se redujo en los hogares ente un 27-43%.70 Para los animales de laboratorio, las últimas tablas de requerimientos nutricionales del National Research Council (NRC) publicado en 1995 no especifican las cantidades de PUFA necesarios en la dieta,71 pero se sabe que es importante su administración para evitar una deficiencia de ácidos grasos que provoca signos como: dermatitis, hígado graso, baja de peso y problemas reproductivos.72 Las recomendaciones alimentarias van cambiando conforme a nuevos descubrimientos en el área nutricional.68,69
Finalmente, los estudios realizados ex vivo/in vitro no reflejan completamente la complejidad de la estructura pulmonar ni las interacciones patógeno-hospedero.73 Los estudios en sangre completa ex vivo tienen la ventaja sobre los cultivos in vitro de permitir evaluar la integración de los efectos de las terapias antimicobacterianas a través de la respuesta inmune del hospedero37 y permiten cada vez acercarnos más a conocer los mecanismos moleculares involucrados en la TB.
Requerimientos de los futuros diseños experimentales
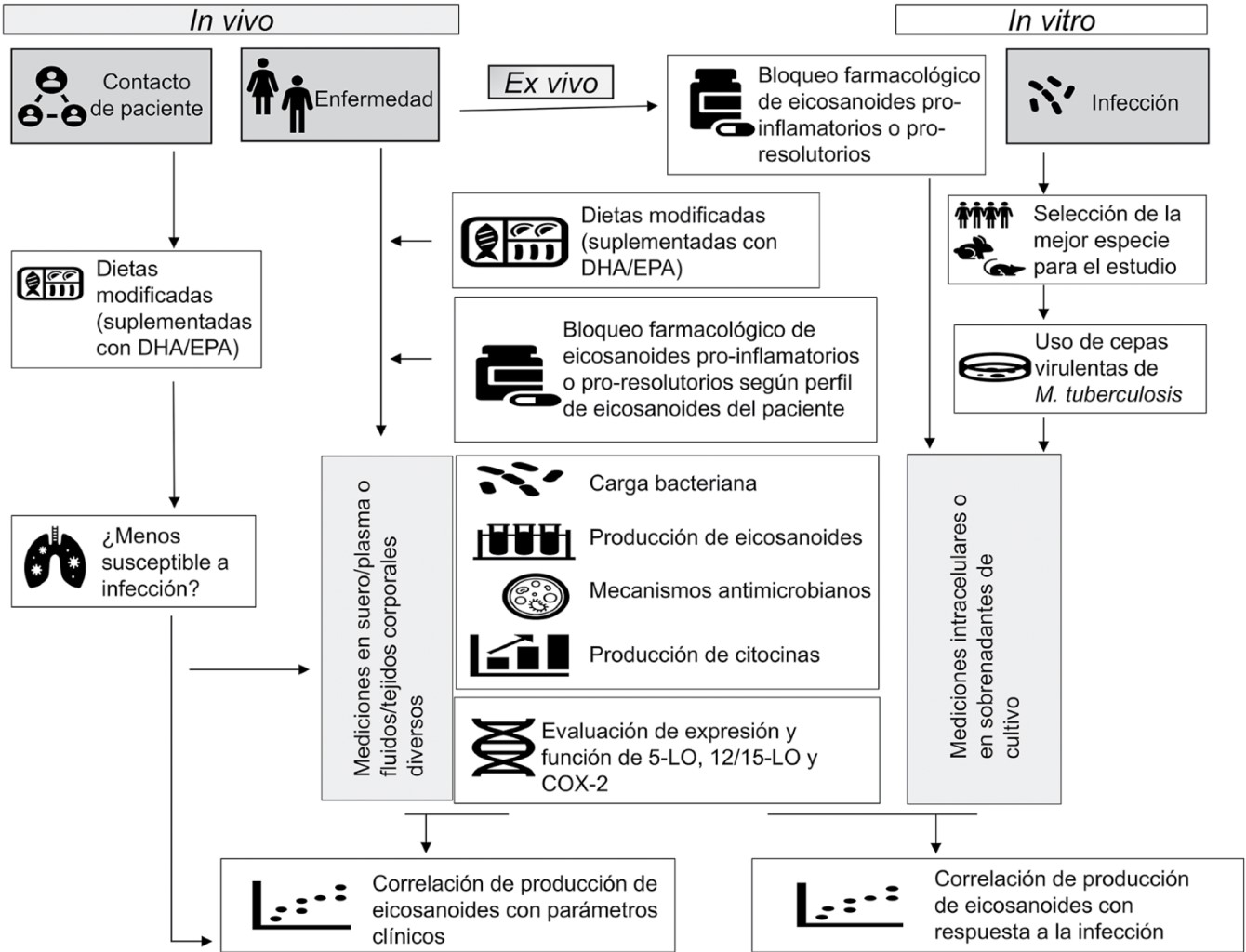
Las intervenciones en la ruta metabólica de los eicosanoides ofrecen diferentes blancos terapéuticos para TB que permiten reducir la inflamación pulmonar para preservar la funcionalidad del pulmón sin pérdida de la inmunidad antimicrobiana. Para que futuras investigaciones permitan conocer mejor el mecanismo de acción de los eicosanoides y proponer esquemas terapéuticos efectivos en TB, se requiere optimizar las estrategias experimentales (Figura 3). Ya sea que se trate de investigaciones in vivo, in vitro y ex vivo, será importante el uso de cepas virulentas de TB para entender mejor las interacciones metabólicas huésped-parásito. Además, es necesario priorizar las investigaciones en humanos, in vitro y ex vivo, tanto pacientes con TB como sus contactos. Para conocer el alcance real de las intervenciones en la ruta de los eicosanoides será necesaria la medición de la carga bacteriana, producción de citocinas y la actividad antimicrobiana celular, así como definir el perfil completo de producción de eicosanoides con miras a medicina personalizada, tomando en cuenta el perfil previo y la fase de la enfermedad en que se encuentre el paciente.
Conclusiones
Debido a que los eicosanoides ofrecen blancos terapéuticos de interés para la TB, es importante optimizar los modelos experimentales y su impacto en la generación de estos blancos. Las dietas con suplementación de DHA/EPA y el bloqueo farmacológico ya sea de eicosanoides proinflamatorios o prorresolutorios podrían ser benéficas tanto para el paciente como el contacto del paciente. Los eicosanoides no sólo tienen funciones en la respuesta inflamatoria, sino que actúan también como mediadores del proceso de patogénesis, por lo que es necesario profundizar la investigación para entender mejor el potencial que tienen los eicosanoides como futuras terapias dirigidas al hospedero.
Agradecimientos
Esmeralda Juárez agradece el apoyo de CONACyT (grant A3-S-35173).
AFILIACIONES
1Universidad Nacional Autónoma de México, Ciudad de México, México 2Instituto Nacional de Enfermedades Respiratorias Ismael Cosío Villegas, Ciudad de México, México.Conflicto de intereses: los autores declaran no tener conflicto de intereses.
REFERENCIAS
Guzmán-Beltrán S, Carreto-Binaghi LE, Carranza C, Torres M, Gonzalez Y, Muñoz-Torrico M, et al. Oxidative stress and inflammatory mediators in exhaled breath condensate of patients with pulmonary tuberculosis. A pilot study with a biomarker perspective. Antioxidants (Basel). 2021;10(10):1572. doi: 10.3390/antiox10101572.
Vinhaes CL, Oliveira-de-Souza D, Silveira-Mattos PS, Nogueira B, Shi R, Wei W, et al. Changes in inflammatory protein and lipid mediator profiles persist after antitubercular treatment of pulmonary and extrapulmonary tuberculosis: A prospective cohort study. Cytokine. 2019;123:154759. doi: 10.1016/j.cyto.2019.154759.
Sorgi CA, Soares EM, Rosada RS, Bitencourt CS, Zoccal KF, Pereira PAT, et al. Eicosanoid pathway on host resistance and inflammation during Mycobacterium tuberculosis infection is comprised by LTB4 reduction but not PGE2 increment. Biochim Biophys Acta Mol Basis Dis. 2020;1866(3):165574. doi: 10.1016/j.bbadis.2019.165574.
Liu S, Xie Y, Luo W, Dou Y, Xiong H, Xiao Z, et al. PE_PGRS31-S100A9 Interaction Promotes Mycobacterial Survival in Macrophages Through the Regulation of NF-κB-TNF-α Signaling and Arachidonic Acid Metabolism. Front Microbiol. 2020;11:845. Available in: https://www.frontiersin.org/article/10.3389/fmicb.2020.00845/full
Peres-Buzalaf C, de Paula L, Frantz FG, Soares EM, Medeiros AI, Peters-Golden M, et al. Control of experimental pulmonary tuberculosis depends more on immunostimulatory leukotrienes than on the absence of immunosuppressive prostaglandins. Prostaglandins Leukot Essent Fatty Acids. 2011;85(2):75-81. doi: 10.1016/j.plefa.2011.04.024.
Rangel Moreno J, Estrada García I, De La Luz García Hernández M, Aguilar Leon D, Marquez R, Hernández Pando R. The role of prostaglandin E2 in the immunopathogenesis of experimental pulmonary tuberculosis. Immunology. 2002;106(2):257-266. Available in: http://doi.wiley.com/10.1046/j.1365-2567.2002.01403.x
Hernández-Pando R, Orozco-Esteves H, Maldonado HA, Aguilar-León D, Vilchis-Landeros MM, Mata-Espinosa DA, et al. A combination of a transforming growth factor-β antagonist and an inhibitor of cyclooxygenase is an effective treatment for murine pulmonary tuberculosis. Clin Exp Immunol. 2006;144(2):264-272. doi: 10.1111/j.1365-2249.2006.03049.x.
Narendran G, Kavitha D, Karunaianantham R, Gil-Santana L, Almeida-Junior JL, Reddy SD, et al. Role of LTA4H polymorphism in tuberculosis-associated immune reconstitution inflammatory syndrome occurrence and clinical severity in patients infected with HIV. PLoS One. 2016;11(9):e0163298. doi: 10.1371/journal.pone.0163298.
Reeves PG, Nielsen FH, Fahey GC. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr. 1993;123(11):1939-1951. Available in: https://academic.oup.com/jn/article/123/11/1939/4722783
Jontvedt Jorgensen M, Nore KG, Aass HCD, Layre E, Nigou J, Mortensen R, et al. Plasma LOX-products and monocyte signaling is reduced by adjunctive cyclooxygenase-2 inhibitor in a phase i clinical trial of tuberculosis patients. Front Cell Infect Microbiol. 2021;11:669623. doi: 10.3389/fcimb.2021.669623.